|
||||
|
|
||||
|
||||
|
|
||||
|
Metabolsimo de las lipoproteínas Dres.
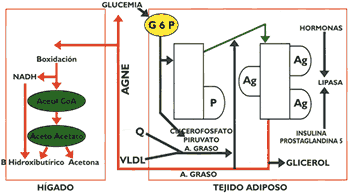
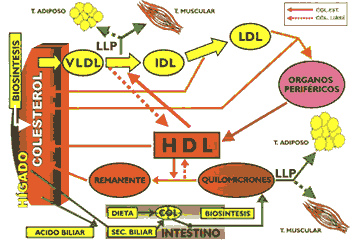
Leonidas Delgado Butrón(*) Introducción Una característica que es común para los lípidos plasmáticos, como el colesterol, fosfolípidos y triglicéridos, es su insolubilidad en el agua, propiedad que los conduce a la necesidad de tener que asociarse con un tipo especial de proteínas denominadas apolipoproteínas o apoproteínas, para ser transportados en un medio eminentemente acuoso, como es el plasma. Las apolipoproteínas son compuestos que poseen una singular propiedad intrínseca que les permite interactuar en interfase, es decir, por una parte de la molécula interacciona con los lípidos y por la otra con la fase acuosa, esta propiedad se manifiesta debido a que termodinámicamente son más estables cuando se sitúan en la superficie de una partícula, lo cual es muy importante para estabilizar la lipoproteína (1). El interés sobre el estudio de las lipoproteínas se tomó creciente cuando a mediados de la década del 50, se describieron técnicas electroforéticas que permitieron separar de una manera relativamente sencilla las lipoproteínas del plasma, cuyos niveles se vinculó con algunas enfermedades cardiovasculares, al mismo tiempo se sugirió que los lípidos que circulaban en la sangre eran transportados por estas lipoproteínas`. Las lipoproteínas no solamente se encargan de transportar los lípidos del organismo, sino que participan activamente en los procesos de regulación metabólica, debido a que las diversas apoproteínas que la conforman interaccionan con receptores celulares de una manera específica, vinculándolas de esta manera con las transformaciones metabólicas que ocurren a nivel intracelular (2); el descubrimiento de los receptores de apolipoproteínas permitió abordar de una forma integral los procesos de regulación lipídica en el ser humano, y conforme decía el Dr. Bruce Campell "Cada apoproteína es una verdadera llave designada a abrirunapuerta específica". La diferente composición y estructura de las lipoproteínas se deben fundamentalmente a su origen y a la naturaleza de las apolipoproteínas que la integran, éstas en cierto modo, son las responsables de la movilidad electroforética de las lipoproteínas y lo mas importante es que el tipo de transformación metabólica de cada una de ellas depende fundamentalmente de la clase de apolipoproteína, por cuyo motivo, una modificación en su composición puede ser la causa de un metabolismo anormal y por consiguiente de ciertas patologías `. Las lipoproteínas difieren en razón a su composición, estructura, densidad y carga eléctrica, propiedades que han inducido al uso de diversas técnicas con el propósito de separarlas, la utilización de la centrifugación analítica permite identificar cuatro clases de lipoproteínas de densidad diferente, en cambio, la electroforesis hace lo propio aprovechando la carga eléctrica que caracteriza a cada una de ellas. Estas lipoproteínas cuyas características se muestran en la Tabla # 1 son: quilomicrones, caracterizados por ser las lipoproteínas de mayor tamaño y menor densidad, su presencia en el plasma, torna a éste de un aspecto latescente; tiene un alto contenido de triglicéridos, que son de origen exógeno, y un pequeño porcentaje de apoproteínas (4). Las VLDL (del inglés very low density lipoprotein) llamada también lipoproteína de muy baja densidad, o pre B- lipoproteína a causa de su movilidad electroforética, es de un tamaño menor que los quilomicrones y en su composición predominan los triglicéridos, que son de origen endógeno, también contiene una apreciable cantidad de colesterol (7). Las LDL (low density lípoprotein) o lipoproteína de baja densidad, también se le conoce como B-lipoproteína, se caracteriza por ser la lipoproteína con un mayor contenido de colesterol y escasos triglicéridos, la apolipoproteína predominante es la apo P-100 (6). Las HDL (hígh density lipoprotein) o lipoproteína de alta densidad, llamada también a-lipoproteína por su movilidad electroforética, es la lipoproteína de menor tamaño siendo sus principales apolipoproteínas las apo A-I y A-II (8). Los ácidos grasos no esterificados, conocidos también como ácidos grasos libres, circulan en la sangre ligados a la albúmina asociación que no constituye propiamente una lipoproteína. Su presencia en el plasma se debe a la lipolisis que ocurre en el tejido adiposo como consecuencia del efecto de las hormonas lipolizantes como el glucagon, que a través de un segundo mensajero como el AMPC activa a la triglicérido lipasa sensible a la acción hormonal (Gráfico 1) (3).
Una precisa descripción de la estructura de una lipoproteína es aquella que corresponde a las HDL3; esta partícula lipoproteica tiene forma esférica encontrándose los triglicéridos y el colesterol esterificado localizados en el núcleo de la lipoproteína, mientras que rodeando a estas moléculas y disponiendo parte de su estructura hacia la superficie se encontrarían los fosfolípidos y el colesterol libre, en cambio las apolipoproteínas estarían ubicadas en la superficie de la HDL3 (8). QuilomicronesSon lipoproteínas que se estructuran en el intestino, utilizando los lípidos que se han ingerido con la dieta. Entre los componentes de esta lipoproteína se encuentran los triglicéridos que se han sintetizado en el enterocito a partir de los productos de hidrólisis de los triglicéridos de la dieta, los fosfolípidos y el colesterol, este último compuesto no solamente corresponde al que se ha ingerido con los alimentos sino a aquel que se secreta con las bilis y al colesterol que se sintetiza "de novo" en el enterocito. Tanto el colesterol dietario como el que acompaña a la secreción biliar, requieren para su absorción de la participación de las sales biliares conjugadas, compuestos que básicamente cumplen dos funciones durante la digestión y absorción de los lípidos, en primer lugar emulsifican a las grasas, con el fin de incrementar la superficie de contacto entre los lípidos y las enzimas encargadas de hidrolizarlos, y en segundo lugar, estructuradas formando micelas simples, las que se encargan de transportar los productos de la hidrólisis enzimática, tal como el colesterol libre, los monoacilgliceroles, ácidos grasos no esterificados, etc. hasta las microvellocidades intestinales donde son posteriormente absorbidos ". Por otro lado, las sales biliares se absorben a nivel del ileon y por la vena porta llegan al hígado para ulteriormente ser nuevamente secretadas al tracto digestivo, formándose de esta manera lo que se conoce como "circulación enterohepática de las sales biliares"; el conocimiento de este proceso, constituye el fundamento del tratamiento de ciertas hipercolesterolemias que ha conllevado al uso de resinas de intercambio iónico, tal como la colesteramina, compuesto que tiene la propiedad de interactuar electrostáticamentecon la carga negativa de las sales biliares impidiendo, de este modo, que éstas se absorban a nivel intestinal, efecto que trae consigo la interrupción de la circulación enterohepática y como consecuencia de ello el hígado se ve forzado a utilizar colesterol para sintetizar sales biliares y restituir aquellas que se han perdido por las heces (16).
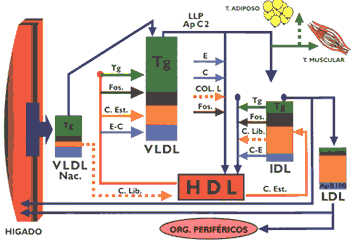
Las principales apolipoproteínas que integran los quilomicrones, se observa en la Tabla 1. Esta lipoproteína se estructura principalmente con los triglicéridos que han sido resintetizados a nivel intestinal, los que se asocian con las apo A-I, A-II y fundamentalmente con la apo B-48 para originar los quilomicrones nacientes que por la vía linfática llegan a la circulación general donde reciben apolipoproteínas que le servirán para metabolizarse posteriormente, como son las apo C- C-II, C-III y apo E cedidos por las HDL (4) (7). Los quilomicrones se dirigen principalmente al tejido adiposo y secundariamente al tejido muscular donde sufren la acción hidrolítica de la lipasa lipoproteica, enzima localizada en el endotelio capilar cuya síntesis es regulada por la insulina y requiere para su actividad la presencia de la apo C-II. Los ácidos grasos uno de los productos de hidrólisis penetran en el adipocito donde servirán para sintetizar triglicéridos, para cuyo propósito es necesario que la glucosa penetre en esta célula y se degrade por la vía glicolítica para proporcionar el glicerol-3-fosfato, componente indispensable en la formación de los triglicéridos(15). El glicerol, otro de los productos formados por acción de la lipasa lipoproteica, no ingresa al adipocito, dirigiéndose al hígado para su posterior transformación metabólica (Gráfico 1 y Gráfico 2).
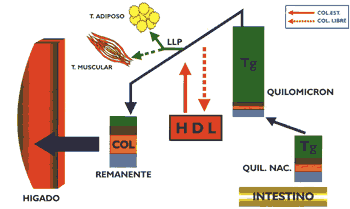
Los quilomicrones, a causa de la acción hidrolítica de la lipasa lipoproteica, disminuirán de tamaño y al mismo tiempo perderán paulatinamente las diversas apolipoproteínas, manteniendo entre otras a la apo E, así mismo, se desprenderán pequeñas partículas discoidales, que originarán las HDL nacientes. Al mismo tiempo participará en una reacción de intercambio con las HDL proporcionando a ésta colesterol libre, la que a cambio le proporciona colesterol esterificado. Esta serie de transformaciones originan una partícula de tamaño límite denominada "remanente" de quilomicrón en cuya composición se encuentra la apo E que les permitirá ser captado por receptores de apo B/E y receptores de apo E que existen en el hígado, lugar donde los remanentes son internalizados para ser catabolizados (4) (19) ; en este proceso la apo B-48 no es reconocida por el receptor antes citado (Gráfico 3). Es necesario considerar que estas partículas no solamente transportan al hígado el colesterol dietético, sino aquel que le ha sido proporcionado por las HDL, contribuyendo de esta manera a su eliminación por la vía biliar. Igualmente, es necesario tener en consideración que los "remanentes" circulan durante un tiempo relativamente prolongado en el plasma, por cuyo motivo, para la determinación del perfil lipídico es necesario que el paciente guarde un ayuno de 13 horas, condición en la que normalmente no deberá encontrarse quilomicrones circulantes (6).
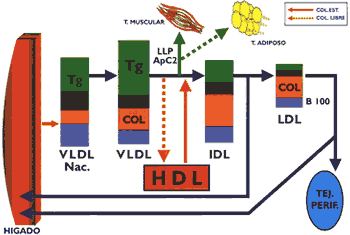
Las pre-betalipoproteínas se estructuran predominantemente en el hígado, siendo la contribución del intestino muy discreta. El mayor componente de las VLDL son los triglicéridos, que se sintetizan a partir de metabolitos proporcionados por el metabolismo de la glucosa, tal como el ATP, NADPH y acetil-CoA, que sirven no solamente para sintetizar ácidos grasos, sino para la síntesis del colesterol. La estructuración de las VLDL depende fundamentalmente de la presencia de triglicéridos, cuya disponibilidad constituye el factor que regula la secreción de la apolipoproteína B-100 componente imprescindible para las VLDL (Gráfico 4) (15). Los triglicéridos sintetizados a partir de los ácidos grasos liberados por el tejido adiposo durante la inanición, no inducen una apropiada secreción de apolipoproteína B-100, lo que trae consigo una acumulación anormal de triglicéridos en el hígado produciéndose el llamado hígado graso. Los fosfolípidos, el colesterol libre y el colesterol esterificado, principalmente estos dos últimos, parecen influir en la secreción de apo B-100 (9). Es necesario precisar que los genes que codifican a las apo B-48 y apo B- 100 en el intestino e hígado respectivamente tienen la misma composición, pero resulta que una modificación post-transcripcional en el enterocito convierte un triplete en el RNAm que codifica el aminoácído glutamina en una señal de parada, lo que ocasiona que la síntesis de esta apolipoproteína se interrumpa cuando se ha traducido aproximadamente la mitad del mensaje (12).
La estructuración de las VLDL se realiza a través de una asociación secuencial de sus componentes, en primer lugar las apo B-100 se asocian a los fosfolípidos y triglicéridos originando una partícula rica en triglicéridos, posteriormente se unen al resto de fosfolípidos y colesterol. Una fase importante del proceso lo constituye la glicosilación de las apolipoproteínas, la que se inicia en el retículo endoplásmico y culmina en el aparato de Golgi. En esta etapa juega un rol muy importante la proteína microsomal transferidora de triglicéridos (MTP), quc interactúa con las apo B- para ensamblar y secretar las VLDL. Luego, éstas se concentran en las vesículas secretoras y se dirigen a la membrana basal donde se fusionan y por un mecanismo de exocitosis se liberan al espacio de Disse, estas VLDL, nacientes están constituidas básicamente por apo B-100. Cuando las VLDL nacientes llegan a la circulación general ceden su colesterol libre a las HDL y captan de esta lipoproteína el colesterol esterificado y las apolipoproteínas C y E. Las VLDL se catabolizan preferencialmente en el tejido muscular, donde se encuentra la lipasa lipoproteica, enzima que también está presente en el endotelio capilar del tejido adiposo, glándula mamaria, y otros, ésta es activada por la apo C-11 e hidroliza a los triglicéridos, liberando ácidos grasos no esterificados y glicerol, de una manera similar al proceso metabólico que ocurre en los quilomicrones (1) (4) (13) . La actividad de la lipasa lipoproteica depende de la relación de las apo C-III/C-II, que se encuentran en la superficie de las VLDL, cuando esta relación se incremento disminuye el catabolismo de esta lipoproteína, conforme se ha observado experimentalmente. Como consecuencia de la acción hidrolítica de esta enzima las VLDL disminuyen de tamaño, de una manera similar a lo ocurrido con los quilomicrones, originándose las lipoproteínas de densidad intermedia (lDL), partículas que pueden ser captadas por los receptores de apo B/E o apo E que se encuentran en el hígado para luego ser intemalizadas, o en su defecto pueden convertirse en LDL, para cuyo propósito es necesaria la participación de la lipasa hepática (2)(19) (Gráfico 5)
Después de una ingesta alimenticia, la liberación de insulina produce un incremento de lipasa lipoproteica, mientras que la fipasa sensible a la acción hormonal, que se encuentra en el interior del adipocito, disminuye su actividad. En cambio, en la etapa post-absortiva, disminuyen los niveles de insulina y aumentan los de glucagon, hormona que activa la adenil ciclasa del adipocito y posteriormente la actividad de la lipasa sensible a la acción hormonal, liberándose como consecuencia de ello ácidos grasos no esterificados que posteriormente se dirigen al hígado (3). Algunos procesos vinculados con bajos niveles de insulina producen una disminuida liberación de lipasa lipoproteica en el tejido adiposo, efecto que también se ha observado en el tejido muscular. Esto trae consigo un limitado catabolismo de las VLDL incrementándose su concentración en el plasma, pudiéndose observar una hipertrigliceridemia que puede revertirse mediante la administración de insulina o hipoglicemiantes orales (14). Se ha observado en aquellas personas que ingieren una dicta muy alta en carbohidratos simples, o en pacientes diabéticos no controlados, la secreción de una VLDL- rica en triglicéridos, la que también se ha descrito en obesos y pacientes con hipertrigliceridemia familiar. Estas VLDL-rica en triglicéridos son retiradas del plasma sin que previamente se conviertan en LDL, proceso en el cual jugaría un rol determinante la apo E, también se ha postulado la participación de la proteína transferidora de ésteres de colesterol (CETP), que tiene la propiedad de transferir triglicéridos a la HDL o LDL, y hacer lo propio con los ésteres de colesterol que son transportados de estas partículas a las VLDL-rica en triglicérido, de tal manera que ésta queda convertida en una lipoproteína remanente enriquecida de apo E, que no se convierte en LDL y puede ser directamente eliminada de la circulación (16)(19). Un aspecto que es importante considerar es el polimorfismo que presentan las apo E, las cuales tienen 3 formas alélicas: E2, E3 y E4, que se diferencian en razón a los aminoácidos cisteína y arginina. El fenotipo E3/3 se presenta en aproximadamente el 60% de la población, una mutación en esta apolipoproteína puede causar dislipidernias debido a un metabolismo anormal de las VLDL. La presencia de las apo E2 y E4 se (12)(13) encuentran vinculadas a ciertas dislipidemias. Lipoproteínas de baja densidad.
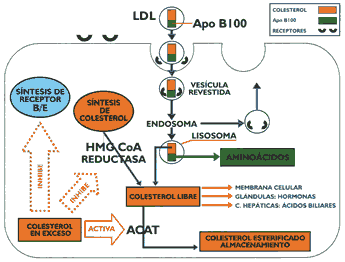
Las LDL se forman a partir de las VLDL y están constituidas básicamente por colesterol esterificado, siendo la apo B-100 su principal apolipoproteína (Gráfico 6). Estas lipoproteínas transportan el mayor porcentaje de colesterol en el organismo y existen varios tipos los que difieren poi su diverso contenido en lípidos y proteínas, constituyendo su elevada presencia en sangre un factor de riesgo positivo para aterosclerosis. Gracias a la presencia de la apo B- 100 la LDI, es captada por órganos como el hígado que posee receptores de apo B/E, así como, por el intestino y los órganos que sintetizan hormonas esteroides y sexuales. Existe también una captación de LDL que no está mediada por receptores, y finalmente, las células macrófagas de Küpffer que poseen receptores con capacidad para reconocer LDI, modificadas (1) (13). Los monocitos y macrófagos a nivel del endotelio vascular tienen la capacidad para interaccionar a través de receptores no saturables con las LDL oxidadas, proceso que a diferencia de los receptores de apo B/E, carecen de mecanismos de regulación, de tal manera que los macrófagos pueden captar las LDL sin limitación alguna, esta interacción que ocurre a nivel vascular conduce a la formación de las "células espumosas" como un evento previo a la formación de la estría grasa (18). Las LDL oxidadas también promueven la secreción de citokinas por las células endoteliales y deprime la liberación de óxido nítrico que es un potente vaso dilatador. La interacción de la apolipoproteína (a) con la apo B100 de las LDL forma la lipoproteína (a) (11), partícula que tiene la propiedad de inhibir el factor beta transformador del crecimiento (TGF-B) cuya función es frenar la proliferación muscular lisa, de tal manera que al inhibirse ocasiona una hipertrofia (4)(10) que es característica de la placa ateromatosa
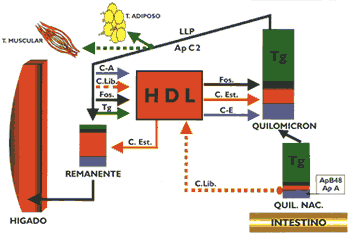
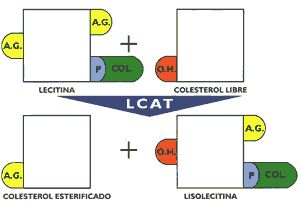
Los receptores de apo B/E, que principalmente se encuentran en el hígado, están dispuestos en depresiones de la membrana celular denominados "hoyos revestidos" recubiertos con una proteína denominada clatrina (Gráfico 7). Cuando interacciona la LDL con el receptor se estructura una vesícula con la formación posterior de un endosoma que se fusiona con lisosomas, cuyas enzimas hidrolíticas degradan la apo B- 100 y a los ésteres de colesterol El colesterol libre formado puede ser utilizado para integrarse a las membranas celulares, activar a la acil CoA colesterol acil transferasa enzima encargada de esterificar el colesterol a fin de que permanezca almacenado, limitar la síntesis de colesterol para cuyo propósito inhibe a la beta-hidroxi beta-metil glutaril CoA reductasa, enzima reguladora de la mencionada síntesis, o inhibir la síntesis del receptor de apo B/E, todas estas reacciones permiten regular el contenido del colesterol intracelular y por extensión el colesterol circulante (13) (15) (16). La incapacidad de una célula para sintetizar receptores de apo B/E conduce a una severa hipercolesterolemia, provocando que los pacientes rara vez superen la segunda década de su vida, debido a su muerte precoz por infarto de miocardio. Se han descrito múltiples mutaciones en el locus del receptor que ocasiona esta patología, existen cuatro clases de defectos moleculares que conducen a la síntesis de receptores de apo B/E defectuosos o ausentes, el primero de ellos, el mas frecuente, muestra incapacidad manifiesta para sintetizar el receptor. Existen mutaciones que dificultan el normal transporte de los receptores a la superficie de la célula, otras mutaciones producen receptores defectuosos y finalmente existen mutaciones que permiten expresar un receptor que ligue normalmente a las lipoproteínas pero incapaces de internalizarla (2)(10)(13). La glucosa normalmente puede reaccionar con las proteínas sin intervención de enzimas formando compuestos covalentes denominados "productos de Amadori", los que a través de reacciones posteriores originan los "productos finales de glicosilación avanzada" (AGE). Este proceso se realiza de una manera más intensa en la diabetes mellitus acumulándose los AGE eneltejido conectivo de las arterias, de tal manera que cuando las LDL se depositan en la pared arterial se impide su difusión promoviéndose la formación de AGELDL, partícula que es reconocida y captada por los macrófagos, que conforme se ha descrito previamente promueve la formación de la placa ateromatosa (17). Lipoproteínas de alta densidadLas lipoproteínas de alta densidad se caracterizan por ser las mas pequeñas y mostrar cierta heterogeneidad en razón a la clase y proporción de los lípidos que la integran, cuyo porcentaje en relación con las apolipoproteínas no es tan alto como en las lipoproteínas previamente descritas. Sus principales apolipoproteínas son las apo A-1 y A-II Esta lipoproteína, tiene diversos orígenes secretándose en el hígado e intestino como partículas discoidales denominadas genéricamente HDL nacientes. Estas están constituidas por las apolipoproteínas A, lecitinas y en menor porcentaje colesterol y triglicéridos. Se ha evidenciado la formación de partículas similares a las HDL nacientes a partir de los quilomicrones y de las VLDL, proceso que ocurre durante el metabolismo de éstas. Las HDL nacientes se dirigen a tejidos extrahepáticos donde captan el colesterol libre que se dispone en su superficie, lugar en que se encuentran también moléculas de lecitina y las apolipoproteínas antes mencionadas. Existe una enzima circulante activada por la apo A-1 denominada lecitina colesterol acil transferasa (LCAT), que tiene la propiedad de esterificar el colesterol con un ácido graso del carbono 2 de la lecitina, la que al perder dicho ácido graso queda convertida en lisolecitina condición en que abandona la HDL para posteriormente esterificarse con un ácido graso no saturado (Gráfico 8). Mientras que, el colesterol esterificado que se ha formado penetra en el núcleo de la partícula, la que incrementa su volumen, después de producirse un gran número de esterificaciones la lipoproteína queda transformada en una partícula madura de forma esférica denominada HDL3 que es la más pequeña de las lipoproteínas circulantes. Posteriormente, ésta incrementa su volumen captando apolipoproteínas en su superficie, así como fosfolípidos y triglicéridos, proceso que la transforma en HDL, cuyos posteriores cambios metabólicos son diferentes y más complejos que el precedente (13)(15)(16). Se ha observado en animales de experimentación alimentados con dietas altas en colesterol, que las HDL, incorporan en su superficie la apo E, así como ésteres de colesterol, originando de esta manera a la HDLc , que tiene algunas características semejantes a las LDL, por cuyo motivo pueden ser captadas por el hígado gracias a la presencia de su apo E (6)(13). Así mismo, se ha descrito un receptor B 1, el que al parecer estaría implicado en la transferencia a los tejidos de los ésteres de colesterol, contenidos en las HDL (5).
La HDL, cede los ésteres de colesterol a las VLDL y a los quilomicrones con intervención de proteínas transferidoras de lípidos neutros y al mismo tiempo capta triglicéridos de estas lipoproteínas, posteriormente con intervención de la lipasa hepática y la lipasa lipoproteica, hidrolizan los triglicéridos y fosfolípidos quedando convertida nuevamente en HDL, Conforme se ha descrito, las HDL son partícipes de un proceso metabólico de tipo cíclico, ya que las HDL, a través de las reacciones descritas se convierten en HDL, la que ulteriormente se transforma en HDL, este conjunto de reacciones en que el colesterol es transportado de los tejidos extrahepáticos al hígado se conoce como "transporte reverso del colesterol" y ello naturalmente tiene sus implicancias en patologías como la aterosclerosis, en cuyo contexto un elevado nivel de HDL sérico constituiría un medio de protección, considerándosele, por tal motivo, un factor de riesgo negativo para aterosclerosis (10,13,16). Adicionalmente, se ha mostrado que las HDL ejercen un efecto protector ya que tienen la propiedad de ligar fuertemente a la paraoxonasa, enzima que degrada lípidos peroxidados afectando probablemente la oxidación de las LDL, que conforme se ha sugerido, estaría vinculado a la aterogénesis (20). El estudio de las lipoproteínas se encuentra actualmente en pleno desarrollo a nivel internacional quedando aún muchos aspectos de su metabolismo por explicar, pero se tiene un apropiado conocimiento de los roles importantes que cumplen en el organismo, especialmente en la fisiopatología de ciertas enfermedades metabólicas cardiovasculares, lo que está permitiendo controlarlas en alguna medida. Resumen 1. El transporte de los lípidos en la sangre lo realizan cuatro clases de lipoproteínas: quilomierones, encargados de transportar lípidos de origen exógeno absorbidos a nivel intestinal; las VLDL que transporta lípidos de origen endógeno sintetizados en el hígado; la LDL que es el producto final del metabolismo de las VLDL, caracterizada por ser la lipoproteína mas rica en colesterol, compuesto que lo distribuye al hígado y tejidos extrahepáticos; las HDL que poseen un alto contenido de apolipoproteínas, fosfolípidos y colesterol, y tienen como función remover el colesterol de los tejidos extrahepáticos e intercambiarlos con otras lipoproteínas para ser conducidas al hígado. 2. La biosíntesis de ácidos grasos, triglicéridos y colesterol hepático se realiza fundamentalmente utilizando compuestos derivados del metabolismo de la glucosa: Acetil CoA, ATP, NADPH y glicerol-3-fosfato. 3. En situaciones de ausencia o escasa absorción intestinal de colesterol, se incrementa la biosíntesis hepática de este compuesto, proceso que es posible inhibir, bloqueando la actividad de la B-hidroxi B-metil glutaril CoA reductasa, enzima que es clave para la síntesis de colesterol. 4. Las apolipoproteínas constituyen los elementos fundamentales para estructurar las lipoproteínas, logrando de esta manera el transporte de los lípidos en un medio acuoso como es el plasma, así mismo, interactúa con receptores específicos, lo que posibilita a una lipoproteína penetrar en una célula. Alteraciones tanto en los receptores como en las apolipoproteínas podrían ocasionar diversas patologías. 5.La enzima lipasa lipoproteica activada por la apo C-II hidroliza los triglicéridos contenidos en los quilomicrones y VLDL, a consecuencia de ello, el primero queda convertido en "remanente" mientras que las VLDL se transforman en IDL, una parte de éstas se cataboliza en el hígado mientras que la fracción restante se convierte en LDL, partícula lipoproteica que tiene en su superficie apo B- 100, por cuyo motivo pueden ser captadas por el hígado y tejidos extrahepáticos por el receptor de apo B/E. 6.Las sales biliares son indispensables para una adecuada absorción de los lípidos a nivel intestinal, su reabsorción y ulterior secreción constituye la denominada "circulación enterohepática de las sales biliares". El conocimiento de este proceso permite en clínica utilizar resinas de intercambio iónico para el tratamiento de ciertas hipercolesterolemias; estas resinas al ligarse a las sales biliares impide que se absorban hecho que obliga al hígado a utilizar colesterol para sintetizar sales biliares con el fin de restituir a aquellas que se han perdido por las heces. 7.Una deficiencia de lecitina o apolipoproteínas a nivel hepático ocasionará una disminuida secreción de VLDL, no obstante que ocurra síntesis de triglicéridos, situación que conduce a un acúmulo anormal de lípidos en el hígado. 8.Los ácidos grasos no esteríficados liberados del tejido adiposo por acción hidrolítica de la triglicérido lipasa sensible a la acción hormonal, son conducidos por la albúmina al hígado, este efecto ocurre por acción de hormonas lipolizantes como el glucagon y ausencia de hormonas como la insulina.
Bibliografía 1. Brewer HB. et al. Apolipoproteins and lipoproteins in human plasma: An overvew. Clin. Chem. 1988; 34:134. 2. Brown MS, Golstein JL. How LDL receptors influence cholesterol and atherosclerosis. Sci. Am. 1994;251:58-66 3. Goldberg W. Lipoprotein lipase and lipolysis: Central roles in lipoprotein metabolism and atherosclerosis. J. Lipid. Res. 1996; 37:693-707. 4. Gotto AMJr, Ponnall HJ, Havel RJ. Introduction to the plasma lipoproteinS. Methods Enzymol. 1986;128:341. 5. Landschulz KT, Pathak RK, Rigotti A, et al. Regulation of scavenger receptor, class B, type 1, a high density lipoprotein receptor in liver and steroidogenic tissues of the rat. J. Clin, Invest.1996; 98:984-995. 6. Me Namara DJ. Effects offat modified diets on cholesterol and lipoprotein metabolism. Ann. Rev. Nutr. 1987;7:273-290. 7. Gibbons GF. Assambly and secretion of hepatic very low density lipoprotein. Biochem. J. 1990; 268:1-13. 8. Edelstein C, K6zdy FJ, Scanu A, Shen B. Apolipoproteins and the structural organization of plasma lipoproteins: human plasma high density lipoprotein-3. J. Lipid Res. 1979; 20:143153. 9. Vance J. Lipoprotein assembly and secretion by hepatocytes. Annu. Rev. Nutt. 1990; 10:337-356.10.Rohein PS. Atherosclerosis and lipoprotein metabolism: Role ofreverse cholesterol transport. Am. J. Cardiol. 1986;57: 3c-1 0c. 11. Scanu AM, Lawn RM, Berg K. Lipoprotein (a). Ann. Int. Med. 199 1; 115:209-218. 12. Fisher EA. Gene polymorphisms and variability of human apolipoproteins. Amm. Rev. Nutt. 1989;9:139-160 13. Palacios E, Osada J. Función del hígado en el metabolismo de las lipoproteínas. En Hepatología Ed. María Cascales y Juan Rod6s. Madrid 1990pag. 43-67. 14. Delgado L, Gu1ja E, Ayala G, Delgado C. Variaci6n de los lipidos s6ricos en la diabetes mellitus. Bol. Sec. Quirn. del Perfi 1979;XIV:152-158. 15. Murray P, Mayer P, Granner D, Rodwell V. Bioquimica de Harper ed. El Manual Modenno S.A. de C.V. M6xico 1997. 16.Herrera, E. Elementos de Bioquimica.- 1993 De. Interamericana S.A. M6xico. 17. Cerami A, Vlassara H, y Brownlee M. Protein glycosylation and the pathogenesis of atherosclerosis. Metabolism 1985; 34:37-44. 18. Kottke B, Lipid markers for atherosclerosis. Am. J. Cardiol. 1986; 57: llc-17c 19. Ginsberg H, Lipoprotein physiology. Endocrinology and Metabolism Clinics ofNorthAmerica (1998)27:503-519. 20. Heinecke J, y Lusis A, Paraoxonase-gene polymorphisms associated with coronary heart disease: Support for the oxidative damage hypothesis? Am. J. Hum. Genet. 1998;62:2024.
(*)Profesor Emérito de la Universidad Nacional Mayor de San Marcos. (**)Profesor Principal de la Facultad de Medicina de la Universidad Nacional Mayor de San Marcos. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||